Summary
- Previously published estimates of the number of birds and mammals killed by owned cats are probably too large by over one billion animals. We estimated a median of 640 million animals killed in both the US and Canada, whereas it has been suggested that a median of 1933 million animals are killed in the US alone.
- Our estimates suggest that interventions aimed at reducing owned cat predation will therefore directly affect a smaller number of wild animals (~830 animals per $1000) than previously suggested (~5500 animals per $1000)
- A single-message advocacy intervention, such as promoting collars with predation deterrents (e.g. bells, bright colors) on all adopted cats, could have multiple benefits for the welfare of both owned cats and wild animals
- However, cat predation may be a net benefit for wild animal welfare given the probability that this mortality is often compensatory, the large reproductive capacity of small rodents, and the inhumane methods of rodent control currently in use
- The widespread use of rodenticides emerges from this analysis a wild animal welfare issue in need of further research with respect to the numbers of target and nontarget animals negatively impacted
Introduction
Numerous articles have focussed on the estimated number of birds, and to a lesser extent mammals and other animals, killed by cats (e.g., Blancher 2013; Loss et al. 2013). These publications have had large impacts in both the scientific and popular press. Cat predation is presented as either a concern for conservation, or animal welfare, or both, and actions to reduce the number of outdoor cats, ranging from extermination to confinement, have been urged.
Scientific and conservation efforts have focused on the potential impact of cats on bird populations. However, in specific jurisdictions, such as Australia, other endangered vertebrates species are important conservation targets. There is no question that where cats are found in areas without an evolutionary history of similar mesopredators, their effects on native fauna can be devastating. The most extreme cases are those where cats have been introduced to island locations with ground-dwelling birds. However, even in these extreme cases, there are some instances of the benefits of cat predation, through the limitation of populations of other nonnative species such as mice, rats and rabbits on native species of conservation interest (e.g., Karl & Best 1982; Flux 1993).
From an animal welfare point of view, cats are predators and do kill other animals. Given the large predation estimates offered by some authors, it seems natural to focus on reducing these kills rates to increase animal welfare. For example, Rowe (2018) compares various interventions and concludes that permanent confinement of owned cats is the most cost-effective means of reducing wild animal deaths due to cats. However, cats are opportunistic and therefore tend to favour abundant or vulnerable prey items (Fitzgerald & Turner 2000). Overabundant prey species also negatively impact other individuals via territoriality, aggressive behaviour, stealing food, or killing young, or more indirectly through competition for limited resources (Gurevitch et al. 1992 Heske et al 1994, Schoener 1983). When resources are limiting, the strongest competition is actually between members of the same species. Therefore, there is a very real possibility that cats, by killing individuals in these populations, reduce both the total population growth rate and the number of individuals that will die of resource limitation or exposure. Since there are no good data-driven estimates of unowned cat populations in Canada and the US (Appendix - Unowned cats), and since there is a large effort promoting owned cat confinement, we focus on the estimated predation rates of owned cats in order to determine if interventions aimed at this population would have a large beneficial impact on animal welfare.
Previous estimates of total cat predation on birds and mammals
Estimates of the mortality caused by cats have clearly indicated the vast majority of cat predation is due to unowned cats (by which we mean undomesticated or feral cats, semi-domesticated cats, and abandoned domesticated cats), if estimates of both population sizes and predation rates are reasonable. For example, using data from the US and EU Loss et al. (2013) estimate that over two billion birds and twelve billion mammals are killed annually by cats in the US. About 69% of this projected mortality is attributed to unowned cats, with the remainder assigned to owned cats. Unfortunately, the population estimates of unowned cats have very wide ranges and very little data-based support (see Appendix - Unowned cats). The lack of data regarding the unowned cat population is extremely worrying in that it drives these large estimates of wildlife mortality. Moreover, we have the example from Australia where estimates of the unowned cat population, for many years proclaimed to be between 10 and 20 million (Dickman & Denny 2010), may actually be as small as 2 million according to recent analyses (Legge et al. 2017). Because of the lack of data regarding unowned cats, and also because there has been so much conservation effort directed toward promoting permanent confinement of owned cats as a beneficial intervention, we focus on owned cats for the remainder of this report – although, in passing, we note that some of the proposed and implemented interventions associated with unowned cats (e.g., poisoning and shooting) are quite inhumane and simultaneously unlikely to achieve stated aims.
The data and distributions used to generate the predation estimates for owned cats in some cases seems to be poorly chosen. For example, Loss et al. (2013) used literature data from the US and EU to estimate that the base of the range of owned cat predation on birds was between 4.2 and 18.3 birds per year (Table 1 Loss et al. 2013). They modelled the distribution of this data as uniform (i.e., assuming that the probability of the predation of 6 birds per annum was the same as 16 birds). An examination of the data the authors used (Supplementary Table S1) suggests that a uniform distribution is not a reasonable choice (this was independently noted by Wolf 2016), and that a lognormal distribution could be a better fit (see Fig 1).
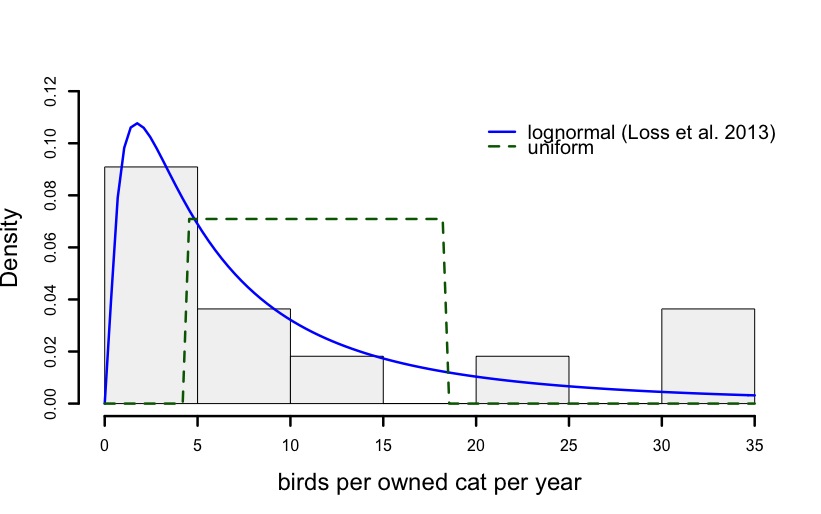
Figure 1: Data from 11 publications for the US and EU used by Loss et al. (2013) to estimate the base predation rate of owned cats on birds with superimposed lognormal and uniform distributions fit using maximum likelihood.
Recently, Loss and coauthors have responded to similar critiques of their choices with vitriolic attacks published in scientific journals (Loss et al. 2018). Here we join with other authors in urging that scientific investigation be continued when there are real uncertainties (Lynn et al. 2019). Contrary to the author’s claims, the model and data choices can have large effects on estimates. Loss et al. (2013) report a median annual bird predation rate of 684 million. Using their original parameters we find very similar results (e.g., median of 685 million, with 95% of the simulations between 258 and 1513 million). If we use a lognormal distribution instead of the uniform distribution to describe the predation rate per cat, then the median estimated number of birds killed by owned cats is reduced by nearly 300 million, although, of course, the interval containing 95% of the simulated estimates is much larger (Fig 2). Understanding how different data and distribution choices affect both the estimated magnitude of predation, and the sensitivity of these estimates to different inputs, is important for relative comparisons between suggested wild animal welfare interventions.
Our estimate of owned cat predation on birds and mammals
As a first step to evaluating the question of whether interventions for owned cats in North America would have a significant impact on bird and animal welfare, we examined the cat population and predation data used by Loss et al. (2013) and Blancher (2013) (see Appendix - Owned cats, Diet and number of prey). We find that for the US there is evidence that the total number of owned outdoor cats has been overestimated, and that this has implications for the estimated number of birds and mammals killed. We also find it likely that the predation rates of owned cats have been overestimated (see Appendix - Diet and number of prey).
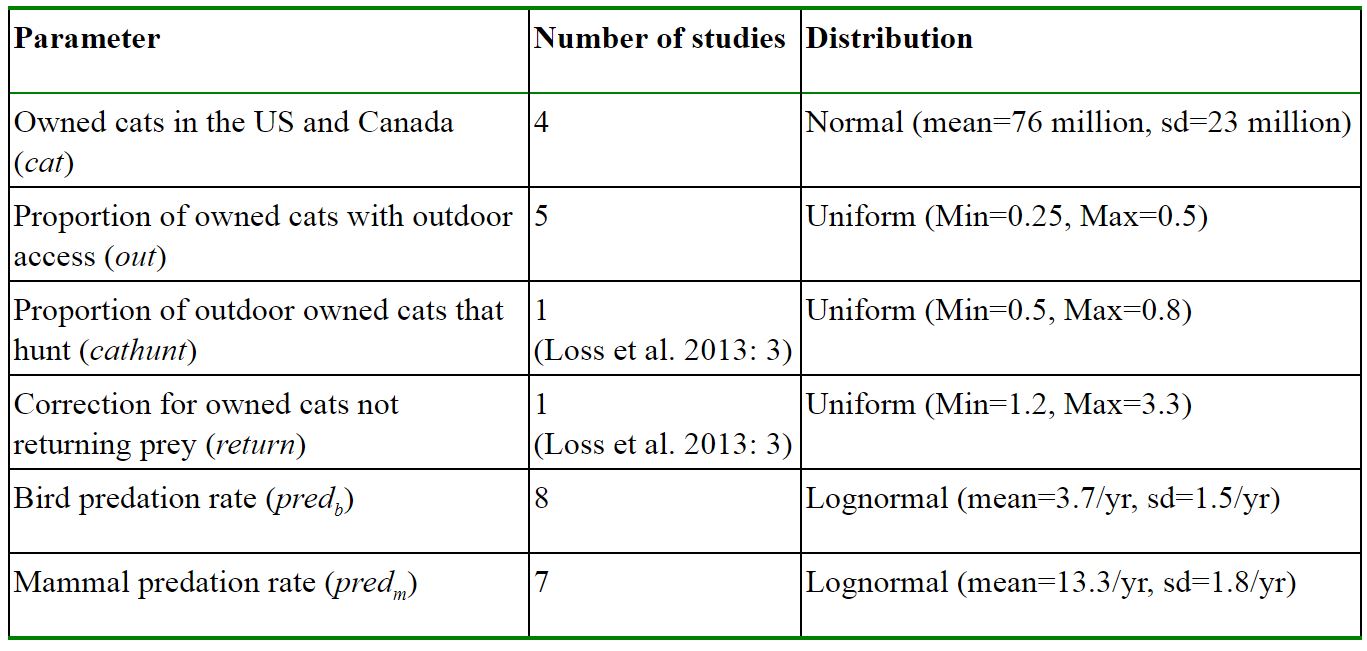
After evaluating the choices made by these authors, we then calculated our own estimated annual mortality rates for birds and mammals in the US and Canada due to owned cats (where kills = # owned cats*percentage outdoors*percentage hunters*bird or mammal predation rate*prey return rate). We note that after vetting only three parameters in the Loss et al. (2013) model – the estimated numbers of owned cats, percentage of outdoor cats, and owned cat predation rates – our estimated median number of annual bird kills is over half a billion individuals lower (see Appendix, Table 1, Fig 2. Code and data at https://github.com/kcudding/ReThinkCats/). This difference is primarily due to the elimination of several questionable references for owned cat predation rates (see Appendix - Predation studies that we excluded).
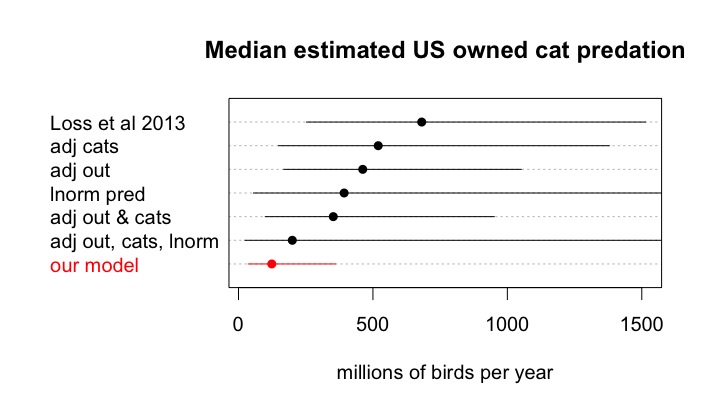
Figure 2: Estimated median number of birds killed (and 95% interval) per year by owned US cats from 10,000 Monte Carlo simulations using either the original parameters and distributions in Loss et al. (2013), the Loss model with cat population numbers adjusted for current data (adj cats), or current percent owned cats allowed outdoors (adj out), or both (adj out & cats), using a lognormal distribution fit to the predation data (lnorm pred), making all three adjustments (adj out, cats, lnorm), or using our model parameters (Table 1). See Appendix for commentary on data sources and parameter values and https://github.com/kcudding/ReThinkCats/ for code.
Using our parameter and distribution choices, we combined the US and Canadian cat population data to estimate the annual number of deaths due to cat predation (see https://github.com/kcudding/ReThinkCats/). We predict a median ~140 million bird kills per year for owned cats in the US and Canada with 95% of the simulation data in the interval between 44 and 397 million. For mammals we predict a median of ~500 million kills per year; however, the interval containing 95% of the estimated values spans orders of magnitude between 128 and 1764 million animals. These estimates suggest that those published by Loss et al. (2013) and Blancher (2013) could overestimate the number of birds killed by owned cats by over 600 million, and the number of mammals by 700 million. Clearly the number of animals killed by owned cats is still large, but for birds especially, these numbers are much smaller than other estimated sources of mortality. For example, Loss et al. (2015) estimate the number of bird deaths due to building strikes in the US at ~600 million and car strikes at ~200 million. In addition, consider that about 9 billion chickens are raised in captivity and killed each year in the US (FAO 2019). The number of mammal kills is much larger, but has very large uncertainty. This smaller estimate of animals killed by owned cats has implications for the cost-effectiveness of interventions such as permanent confinement.
Table 1: Parameter values used to generate our estimate of annual numbers of birds and mammals killed by outdoor owned cats in Canada and the United States (see Appendix).
Evaluation of interventions to reduce owned cat predation
A previous estimate of the number of animal lives saved per $1000 spent on adoption level advocacy urging new owners to permanently confine their cats indoors concluded that a median of 5500 wild animals would be saved (with 95% of the simulations in the range of 2400 to 22,000) (https://www.getguesstimate.com/models/11008). The authors used total kill rates from Loss et al. (2013) to reach this conclusion, but used a smaller range of the percentage of outdoor cats than we did. Using our numbers (Table 1, with constants for reptiles and amphibians from this model added, since we do not have robust data for these), that included a larger range for the percentage of outdoor cats, and the total estimated owned cat population for both US and Canada, we arrive at a median of ~830 animal lives not subject to cat predation (or 0.83 animal lives affected per dollar), with 95% of the simulations in the range between 330 and 2150. This calculation includes the assumption of ~33 cats, which would otherwise have been allowed outdoors, being kept indoors permanently as a result of the $1000 worth of advocacy. Therefore, this intervention may lie within the range of cost effectiveness for corporate campaigns aimed at farmed animals (Šimčikas 2019). Of course, the comparison here is between number of farmed animal lives affected by welfare changes and wild animals not killed. In addition, we must also consider the reproductive efforts of the prey species in order to fully understand the impact (see Welfare impacts below). Unfortunately, comparisons to interventions aimed at unowned cats are beyond the scope of this article, given the extremely large uncertainty surrounding the size of this population.
Following the approach of Loss et al. (2013) and Blancher (2013), we estimated the percent variance explained by each model parameter using the sum of squares from a multiple linear regression (Fig 3). The predation rate was the most important quantity, followed by the estimated number of cats, the correction factor for the prey return rates, the proportion of outdoor cats, and the proportion of cats that are hunters. Sensitivity, as indicated by the standardized regression coefficients, follows the same pattern. A 10% decrease in the percent of outdoor cats (i.e., changing the uniform distribution range to 0.225 to 0.475) decreases the median estimated number of bird kills by ~10 million and that of mammals by ~43 million. In comparison, a 10% decrease in the predation rate (changing mean from 3.7 birds/yr to 3.4, and 13.3 mammals/yr to 12.0 mammals/yr) decreases the number of bird and mammal kills by ~15 million and ~53 million respectively.
The predicted number of deaths due to owned cats was most sensitive to predation rate. Therefore, campaigns aimed at convincing owners to use collars with predation deterrents (e.g., bells, bright colours) may result in fewer animal deaths than interventions aimed at confining cats indoors if the interventions have equal costs and efficacy. In the UK, hunting cats wearing a bell returned 34% fewer mammals and 41% fewer birds than those with a plain collar (Nelson et al. 2005). Cats known to be hunters wearing the Birdsbesafe collar cover killed 19 times fewer birds than uncollared cats in the spring, and 3.4 times fewer birds in the fall in New York state and did reduce mammal predation in the fall, but not the spring (Wilson et al. 2015),. However, this product has little impact on mammal predation in Australia (Hall et al 2015). While it is possible that there are negative impacts of quick-release collars with predation deterrents on cats, we could find no reports in the literature.
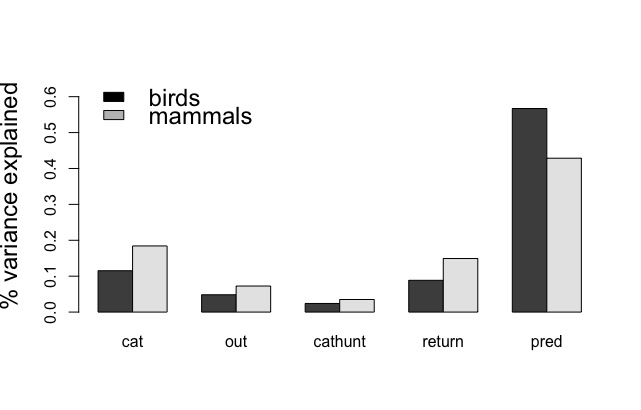
Figure 3: The percent variance in the number of birds or mammals killed explained by each variable in our model of cat predation as given by a multiple linear regression, where cat is the estimated number of owned cats in the US and Canada, out is the percent of these cats allowed outdoors, cathunt is the percent of outdoors owned cats that hunt, return is the correction factor to calculate actual kills from prey return rates, and pred is the predation rate per cat.
It is likely that the costs for an adoption level campaign aimed at collaring cats with predation reduction devices would be similar to the costs estimated for increasing the number of indoor-only cats (Rowe 20188). Such a campaign may even have greater efficacy. In the UK, where permanent confinement is less popular and concern for wildlife lower than other jurisdictions (Hall et al. 2016), collar-mounted predation deterrents were considered the most acceptable intervention (Thomas et al. 2012). However, a complete evaluation would also require an estimate of the percentage of cats that currently wear predation deterrent devices. Wilson et al. (2015) suggest that most cats wear no collar at all, but do not have data on this point. If true, then advocacy on this position could have a large potential impact. Moreover, Lord et al. (2010) indicate that ~73% cats who have not previously worn a collar will tolerate them well. Further, the authors also urge the collaring of indoor cats as a safeguard for identification and recovery in the case of escape, in addition to the use of microchips, since information on collars can be readily accessed. Therefore, a single-message advocacy intervention, such as promoting the use of a belled collar on all adopted cats, could have multiple benefits..
Welfare implications
Before contemplating any intervention, however, it is worth examining which individuals are likely to be targeted by cats in order to understand the welfare implications of this predation. Particularly for mammalian prey, it is unclear whether cat predation is net negative for animal welfare. In addition, there is also the welfare of the cats themselves to consider.
Cats
While American and Canadian veterinary associations recommend indoor confinement as a way to improve welfare, cats that are confined may be subject to physical and mental health issues from the associated inactivity, boredom or stress. For example, Slinglerland et al, (2007) found that the incidence of diabetes was significantly associated with physical inactivity and confinement. This relationship between indoor confinement and diabetes is supported by other studies (Öhlund et al. 2017). The association is almost certainly due to a higher incidence of obesity in confined cats (e.g., Gibson et al. 2018). In addition to diabetes, obesity increases risks of hepatic lipidosis, lameness, oral cavity disease, urinary tract disease, dermatological disease, and neoplasia (Tarkosova et al. 2016). Confinement itself has also been associated with higher incidences of feline urologic syndrome, hyperthyroidism and dental disease (Buffington 2002). In addition, there is a higher incidence of behavioural issues in confined cats (e.g., Amat et al. 2009), which is likely due to increased stress, insufficient stimulation and lack of physical activity (Bain & Stelow 2014). However, cats with outdoor access are more likely to be infected with parasites or pathogens, some of them lethal (Chalkowski et al. 2019). They also engage in risky behaviours outdoors such as crossing roads, entering confined spaces and interacting with other animals that can result in harm (Loyd et al. 2013a). For example, cat deaths under the age of 5 in Britain are most often due to road trauma (McDonald et al 2014). It is difficult to counteract some of the risks experienced by cats allowed outdoors. However, while the negative impacts of confinement can be counteracted by providing a stimulating environment and creating opportunities for exercise, not all owners are willing or able to do so. We conclude that the welfare impact on cats of confining them indoors seems uncertain.
Prey species
The welfare impacts of cats on their prey is not necessarily negative. Cats are opportunistic predators that will target vulnerable prey. For example, Liberg (1984) reports that during a harsh winter in Sweden rabbits were suffering from starvation and cats took a very large number of rabbits that he described as “dead, dying and very weak”. During that winter rabbits made up 93% of all prey, and intake was 6 times higher than in the previous years. Similarly, Baker et al. (2008) report that across species, birds killed by cats had significantly lower mass, fat scores and pectoral muscle mass scores than birds killed by collisions with buildings or automobiles in Bristol UK, even though these birds did not differ in age, wing length or season of death. Therefore, in some cases, cats may selectively target mammals and birds in poor physical condition (Dierschke 2003; Møller & Erritzoe 2000), and they are certainly known to target juvenile animals. These classes of individuals may be expected to have high natural mortality rates in the absence of cats, such that cat predation could wholly or partially compensate for natural mortality rather than being additive to it. In other words, in many cases, cats may preferentially kill individuals that were likely to experience considerable suffering, and then death in the near term anyway.
Birds
As compared to small mammals (see below), the welfare implications of cat predation on birds are particularly unclear, and are probably species-dependent. While some authors have suggested that cat predation or even sublethal effects of predatory behaviour (Beckermann et al. 2007) represent an additive mortality factor in mainland locations, there is little evidence to support this claim. We know that birds killed by cats are more likely to have lower mass, lower fat reserves, and poorer immune systems than other birds (Dierschke 2003; Møller & Erritzoe 2000; Baker et al. (2008)). In addition, cat predation on birds is greatest in the spring, probably reflecting the killing of juvenile individuals (Baker et al. 2005; Lepczyk et al. 2003). However, this is not evidence that these birds would have died a more protracted or painful death in the absence of cats. On the other hand, mere estimates of the magnitude of cat predation or sublethal impacts tell us nothing about whether these animals would have survived in the absence of cats. For example, Malpass et al. (2018) found that while cats were 5x more likely to attack the nests of robins and cardinals in residential neighbourhoods as compared to forest parks of Ohio, there was no difference in the nest survival rates between the two habitats. Even so, it seems plausible that cities and suburban areas with especially high cat densities might expose birds to additive predation. The evidence for this point is unclear. For example, a meta-analysis of reproductive success in urban environments found patterns of lower clutch size, lower nestling weight and lower productivity which the authors attributed to lower levels of natural foods for nestlings (Chamberlain et al. 2009). If true, this implies that nestlings and fledglings in urban environments are more likely to starve than those in more natural surroundings. There is therefore large uncertainty regarding the welfare impacts of cat predation on birds.
Rodents
The small mammalian prey favoured by cats, such as mice and voles, have short lifespans and enormous reproductive capacity, which makes it quite likely that cat predation represents a net benefit to animal welfare for this group. To get some idea of the numbers and astonishing capacity for population growth, consider that Berry (1981) calculated a single pair of house mice could theoretically expand to a population of 2,688 mice in only 6 months, assuming no mortality and a litter size of six young. Gestation is only three weeks, and females are sexually mature at six weeks after birth. However, Berry and Jakobson (1971) estimate that only about 5% of the population will survive a harsh winter (although a mild winter can lead to an outbreak population in the following season). That is, most individuals born are going to die of exposure and starvation within their first year. Therefore, if a cat kills a newly mature female mouse before its first bout of reproduction, over the course of one season it would potentially prevent the addition of over 1000 additional mice to the population (assuming a mortality rate of 50% of the nestlings; Berry and Jakobson (1971)), of which ~950 were going to die over the course of the year (although note that this estimate was made in a nonurban environment; values may differ where abundant human resources are available). Reconsidering the costs of intervention (see above), and assuming 70% of the prey were mammals similar to house mice, we might then conclude that $1000 would potentially affect more than 29 000 mammals, where this figure includes the original 581 mammalian prey, and their potential offspring, assuming a 50% sex ratio. There is considerable uncertainty regarding the central tendency and distribution of welfare levels of different groups of wild animals. In addition, there is some uncertainty about the relative suffering involved in different possible deaths.
Whether or not you view a predation death as a better or worse death than poisoning, disease, starvation or exposure will also depend on how you scale the pain of these events vs. the length of the episode. Cats kill by breaking the neck of their victims so that death is close to instantaneous. In a controlled study, Biben (1979) observed that adult mice are almost always killed by cats with a single bite lasting only a few seconds, while larger prey (young rats) require over a minute until death. The incidence of efficient kills, with quick pursuit and kill bite was correlated with the size of the prey and the hunger status of the cat. With larger prey and lower hunger levels, cats were more likely to engage in play behaviours (Biben 1979). It is hypothesized that this behaviour will exhaust the prey so that injury to the cat is less likely. For the prey, this means that a predation event could take longer, perhaps an hour or so, and moreover, the prey may not be killed, but instead merely fatally injured. For mice, starvation with complete withdrawal of food can take relatively little time (2-3 days), but is probably less likely than severe calorie restriction that takes longer to cause death. However, there is evidence that the chronic pain of starvation is dulled, relative to acute pain responses, in starving mice (Alhadeff et al. 2018). In their discussion, Pocock et al (2004) suggest that most commensal mice in their study died by poisoning and predation, while wild living mice tended to die from exposure, starvation and when population density is high, disease (Berry et al. 1973). Therefore, for rodents likely to be killed by owned cats, the most relevant distinction may be between death by predation vs death by poisoning.
The highest populations of mice, like the highest populations of cats, are generally found near human habitation (Laurie 1946). However, near human habitation, agriculture and industry, rodent populations are subject to control measures such as trapping and poisoning. Rodenticides and kill-trapping methods have associated animal welfare (target and non-target) issues (Mason and Littin 2003). In 2006, the EPA reported that 90% of of the $90 million spent by residents on rodent control was dry bait, a category primarily composed of anticoagulants (U.S. EPA 2006), with the remaining 10% comprising snap traps, glue boards and similar items. For the professional pest control market, 75% to 80% were rodenticide products, with 80% of these being anticoagulents. The remaining 20% to 25% included items such as traps and glue boards (U.S. EPA 2006). As well, the most common self-reported form of rodent control among farmers and pest control agencies is the use of anticoagulant rodenticides (Hindmarch et al 2018, Memmott et al. 2017). Although some residential users in California report they are slightly more likely to use snap traps than anticoagulants, while glue boards are the third most likely control method for this group (e.g., Morzillo & Mertig 2011).
Anticoagulant rodenticides cause a lingering death over days to weeks of severe hemorrhage, with massive internal bleeding and poor coagulation and symptoms associated with blood loss such as anemia, pale mucous membranes, weakness, hypothermia, and tachycardia. Moreover, these rodenticides are known to be the source of secondary poisoning in related predator species (Erickson & Urban 2004), because the poisoned rodents take so long to die, and in their weakened state are an easy target. These chemicals are highly persistent, and accumulate in the tissues of species higher up in the food web (e.g., raptors, mountain lions, fishers), where they cause both lethal and sublethal effects. There is ample evidence for widespread foodweb contamination driven by the use of rodenticides (e.g., Gabriel et al.2018; Regnery et al. 2019; Seljetun et al. 2019; Serieys et al. 2019). In addition, non-target co-occurring rodents, birds, pets or children may ingest the poisoned baits. Unfortunately, new consumer market regulations on the more lethal second generation chemicals do not seem to be having a large impact on overall use or nontarget wildlife contamination (Murray 2017).
The use of rodenticides is related to human occupation and land use, and therefore overlaps those areas where we could have a higher number of outdoor owned cats. A survey of residents in Bakersfield, CA found higher use rates in single-family dwellings next to open spaces, as compared to multi-family dwellings in more urban locations (Morzillo & Schwartz 2011), although is is possible that controls are being implemented by landlords in denser housing and were not detected by this survey. Nogeire et al (2015) used the California agricultural database to classify land-cover types with respect to anticoagulant rodenticide use, as well as the corresponding measurements of exposure of kit foxes in these areas (Cypher et al 2014). Areas with confined animal agriculture, semi-agricultural areas or low-density residential areas (e.g. Morzillo & Mertig 2011) were assigned the highest usage rates. Urban lands and orchards were scored intermediate use while natural land cover, cropland or grazing lands were given a low use score. Outdoor cat densities may be highest in residential areas and industrial areas (Flockhart et al. 2016), and could potentially be exploited to help control rodent populations and reduce the use the use of rodenticides.
It is certainly clear, however, that outdoor cats alone cannot fully control population growth of of small rodents. For example, Liberg (1984) estimated the number of individual field voles and wood mice consumed by cats in an area of 4500 hectares in Sweden over 2 years as ~31 000 voles and ~5000 mice, only about 20% of his estimated production of new voles and mice of 171 000 and 20 000 individuals respectively. However, in sufficient numbers they may exert enough of an influence to make local population increases less likely, and to reduce the need for less humane control methods. For example, some owners keep farm cats in order to control rodent populations (Crowley et al. 2019), or report that cats reduce rodent problems (Hindmarch et al 2018). Aside from predation, outdoor cats may also reduce the rodent populations near human interests by creating a “landscape of fear” (e.g., Themb’alilahlwa et al. 2017). While some studies indicate that cats are not effective predators of larger rodents (e.g., Norway rats Glass et al. 2009), a combination of low levels of predation and the pheromones released by the cats may be enough to repel most species. For example, several animal charities run “cats at work” programs for unowned cats that are not adoptable as pets. These animals are given food, shelter and medical care in return for patrolling areas that are plagued by rodents, and seem to have a high success rate at reducing local population problems (e.g., Washington Post 2016). Such changes in occupation could reduce the use of rodenticides since residential users often report that it is the visibility of rodents that leads them to employ control measures (e.g., Hindmarch et al 2018). Therefore, outdoor owned cats may simultaneously reduce the need for inhumane rodent control near human interests, and reduce fast population growth rates that lead to larger amounts of animal suffering.
Conclusions
We find that the previously published estimates of bird and mammal kills due to owned outdoor cats are probably inflated. As a result, interventions aimed at increasing wild animal welfare via methods to reduce this predation probably have a lower cost-effectiveness than previously calculated. The estimate of deaths due to owned cats was most sensitive to predation rate, suggesting interventions that affect this parameter directly, such as advocating the use of cat collars with predation reduction devices (e.g., bells), may have a larger impact than efforts aimed at keeping cats indoors. Moreover, use of predation reduction devices may be more palatable than permanent confinement to some owners. However, given that the vast majority of previously estimated cat predation (~70%) was attributed to unowned cats, even a 100% effective intervention for owned cat predation would address only a small proportion of wild animal deaths due to cats. Thus research aimed at providing data-based population estimates for unowned cats is sorely needed. This is particularly true since the welfare of unowned cats themselves may also be quite low.
Finally, we note that there is a reasonable possibility that cat predation is actually a net animal welfare benefit. Cats may tend to target animals that have a high probability of a slower death due to poor condition or high juvenile mortality. They may also reduce the need for more inhumane pest control measures in areas of human occupation. In fact, use of rodenticides emerges from this analysis as an animal welfare issue of potentially large magnitude worthy of further investigation. We conclude that it is unclear whether reducing the number of outdoor owned cats would have a net benefit on animal welfare.
Appendix
Current population estimates
The total numbers of cats in Canada and the US may be usefully divided between owned cats and unowned cats (by which we mean undomesticated or feral cats, semi-domesticated cats, and abandoned domesticated cats). Of these populations, only those cats that have access to the outdoors are considered to have a significant impact on rodents and birds for the purposes of this report.
Our review of the literature makes it seem very likely that the estimated numbers of outdoor cats in the US, and possibly those in Canada, used in the conservation literature are overestimated. In a recent contribution, Rowan et al. (2019) and Wolf (2016) similarly argue that there are far fewer outdoor owned cats than these papers suggest. Published papers that include estimates of the number of owned cats in the US normally only cite two sources that are almost certainly inflated. There seems to be only one national survey for Canada. The percentage of owned cats that spend time outdoors has also been estimated on a rather ad hoc basis in most publications, although there is survey data. For unowned cats, previously published estimates are primarily based on expert opinion rather than data, and therefore have large margins of error.
Owned cats
US
Two surveys of pet populations in the US are commonly used to estimate the number of owned cats. The American Veterinary Medical Association (AVMA) conducts a survey every 5 years using representative data (nonrandom) of ~50,000 households, while the American Pet Products Association (APPA) also uses a nonrandom sample and had ~22,000 respondents in 2017. Rowan et al. (2019) report that the AVMA surveys indicate a virtually unchanged rate of cat ownership (around 30–30.5% of households) from 1986 to 2011 while the APPA surveys show a steady increase in the rate of cat ownership (from 30 to 37% over the period from 1988 to 2016).
Currently both surveys use Internet “opt-in” panels, which are much cheaper than randomized mail, telephone or in-person methods. The responses are typically weighted to match U.S. demographics, but this may not be sufficient to produce representative data. Opt-in surveys focused heavily on one topic, such as pet ownership, are more likely to elicit responses from pet owners. The American Association for Public Opinion Research warned that researchers should avoid this method for estimating populations (AAPOR 2010). It is quite likely that APPA survey, with its focus on pet ownership and smaller sample size is a less reliable estimate. Matt Salois from AVAM indicated to the Washington Post (Jan 31, 2019) that they specifically targeted non-pet owning households in their 2016 survey, which had 41,000 respondents. To extrapolate to national numbers, the estimated percentage of cat-owning households are multiplied by the average number of cats owned per household. The APPA survey reports a higher percentage of cat ownership which leads to an estimate of 94 million owned cats in 2016/2017 as compared to the AVMA estimate of 58 million for the same period.
According to the Washington Post (2019), the Simmons National Consumer Study, which is an omnibus survey that occurs annually and which has ~25,000 randomly selected respondents contacted by mail and phone, found 53 percent of households owned pets in 2016, which yields an estimate of 54 million cats, or about the same as the AVMA for the same year. The American Housing Survey from the US Census Bureau is conducted in person from a random sample and has a response rate of 81% from 30,000 households. This survey first asked about pet ownership in 2013 and found that 48% had pets, and 49% 2017. These estimates are both much lower than APPA estimate of 68% in 2017 and 67% in 2019.
Rowan et al. (2019) speculate that even the AVMA numbers may be too high, and provide an example from Massachusetts from 1996 to 2011 that compares survey results from the Massachusetts SPCA and the state level estimates from AVMA. The AVMA cat population estimates are about 50% higher than those of the local agency. In addition, there is considerable variation in cat ownership within and between between states. The more rural states (MT, ND, ME, WY, VT) tend to have higher rates of cat ownership (~45%) while LA, MI, MD and IL had the lowest rates at around 26-28%, so that extrapolation based on nation-wide averages of cat ownership, as done by the APPA survey, may be problematic. As a best guess using the AVMA and Simmons survey, the current number of owned cats in the US is more likely to be in the range of 50-70 million than the 94 million reported by APPA. Taking the mean of these three estimates, we can describe the estimated US population of owned cats as a normal distribution with mean of 67 million and standard deviation of 23 million. Using this estimate with the Loss et al. (2013) model generates a median annual bird predation rate for owned cats that is approximately 170 million lower than the estimate published by these authors..
Canada
A nation-wide survey conducted by Kynetec (formerly Ipsos), estimated the number of owned cats in Canada at 8.3 million in 2018 (CAHI 2019). This number is based on responses of 3,026 pet-owning households, where the percentage of ownership exceeded the highest estimates of US ownership at 37%. These findings are consistent with previous surveys done by Kynetec on behalf of the Canadian Animal Health Institute (CAHI) since 2004. Indeed the survey results are virtually identical to the 2016 online survey by the IPSO with 1,222 respondents. We could find no reported errors about these estimates but Blancher (2013) uses a value of 8.5 million +/- 0.25M in his 2013 study.
US and Canada
Accordingly, we fit a normal distribution to the estimated values of owned cats in the US and Canada, and combined these values. The mean number of US owned cats was given as ~67 million (mean of the two surveys estimating 54 million and the APPA survey estimating 94), for a total North Americanpopulation described as a normal distribution with mean 76 million and standard deviation of 23 million.
Unowned Cats
US
Estimates of the number of unowned cats in the US vary wildly. Moreover, there is little empirical data supporting these estimates. This state of affairs is unfortunate because estimates of the number of birds killed by cats are reported to be quite sensitive to the number of unowned cats (e.g., Blancher 2013). Unowned cat population size explained the greatest variation in prey mortality estimates (42% for birds and 51% for mammals) in Loss et al. (2013), while at the same time 69% of this mortality was attributed to unowned cats. Those authors report in the supplemental material that of the data sources they used “the validity of extrapolating three density values to a national-scale abundance estimate is questionable”.
Loss et al. (2013) note that there are no data-based estimates of the unowned cat population in the US, but use a uniform distribution with minimum of 30 million and maximum of 80 million to define this population in their model. Reports of unowned cat population size that are supported by data range from 10 to 50 million, although estimates without cited data range up to 100 million (e.g., Dauphine & Cooper 2009). Like other authors, Loss et al. (2013) try to extrapolate from local density estimates to obtain a US-wide estimate. For example, Levy et al. (2004) gives a value of 50 million unowned cats vs. an owned cat population of 73 million at that time. Her estimate is based in part on the APPA estimate of owned cats, the percentage of households reporting that they fed unowned cats in four counties from Florida, California, and Massachusetts (~10%) and the household estimate of the number of cats fed (~3.5). The author suggests that unowned cats are a minimum of 36–46% of the population of owned cats, and offers a calculation of 0.5 times the number of households to arrive at her estimate. However, in Maine, a survey conducted Katie Lisnik of The HSUS indicated that the number of unowned cats may be no more than 10% of the pet cat population (cited in Rowan 2018). Rowan (2018) suggests that it is likely that states with hard winters will have far fewer outdoor cats than states like Florida. Merritt Clifton estimates that the unowned cat population ranges from around 6.5 million in winter to around 12.5 million in summer (reported in Rowan 2018).
Loss et al. (2013) attempt to extrapolate by calculating an average density of cats in a local study, and multiply by the area of the US. However, scientific reports from around the world indicate that cat density away from human habitation tends to be low (less than 1–5 cats per square kilometer; Liberg et al. 2000) unless there is some concentrated food source to support higher cat populations. For example, Genovsei et al. (1995) report a stable density of 1.5 cats/km2 in an isolated area of Italy. Hansen et al. (2018) estimated the unowned cat density in a New Zealand national park adjacent to human habitation using camera traps at between 1.39 and 2.58 cats/km2. Worldwide, these estimates are also similar to density estimates for unowned cats with a food source of sparse natural prey (Liberg et al. 2000). In urban centers, these numbers are much higher.
Data-based studies support the claim that cats are most often found in the immediate vicinity of human habitation even in geographically restricted regions. For example, Piquet et al. (2019), in a complete survey of La Graciosa, found that most cats were found near and within anthropic areas (representing 1.63% of the total islet area), whereas the rest of the islet showed a lower cat density. Typically, dense populations of cats occur in locations where resources (e.g. food and shelter) are more concentrated (e.g. Natoli 1985; Liberg et al. 2000).
Glossing over the variance in the spatial distribution of unowned cats can lead to overestimates of both their density and potential impact. The estimation of unowned cat numbers in Australia offers a precautionary example. For some years, it was widely claimed that there were between 10 and 20 million unowned, feral cats on the continent (Dickman & Denny 2010). However, this number was an unsupported estimate (Lynn 2015). When biologists finally undertook a comprehensive assessment of cat numbers in Australia they determined that there were between 1.4 and 5.6 million cats in natural environments (the higher number being present following the rainy season) and a further 0.7 million outdoor unowned cats in highly modified urban and suburban environments, where mean cat densities are “hyper-variable” depending on food subsidy (Legge et al. 2017).
Attempting to use the same methods as Legge et al. (2017), Rowan et al. (2019) estimate that about one quarter of all unowned cats in the USA (~8 million) occur in natural areas in the USA versus 24 million in the relatively small areas of highly modified urban and suburban environments, for a total of approximately 32 million unowned cats. In other words, the vast majority of unowned cats in the US are concentrated in the same urban and suburban areas where humans have highly impacted the environment. Therefore, we concur with Loss et al. (2013), that the number of unowned cats in the US may lie somewhere in the range of 30 million to 75 million, but suspect that it is more likely to be on the low side of this range, or possibly even lower (see below regarding data on unowned cats in one location in Canada).
Canada
In Canada, Blancher (2013) notes that there are no data-based estimates of the unowned cat population, and does incorporate spatial variability in his estimate. He uses media reports from animal care workers compared to the size of the human population in the same municipality to get a range in number of unowned cats per 1000 people of 50 and 150. That ratio was extrapolated to all other municipalities in southern Canada, where southern Canada was defined to exclude 78% of Canada’s area that is in arctic and northern forest areas to give a range of 1.4 to 4.2 million unowned cats in southern Canada.
If we compare this estimate to a local data-based estimate, however, there are large discrepancies. Using distance sighting methods, Hand (2019) estimated unowned cats at about 13.3 (95% CI 9.7–18.1) cats per km2 in Windsor, Ontario, for an estimated population size of 1,858 unowned cats (95% CI 1,361–2,537). Windsor has a human population of ~330,000 which yields a ratio of 4-8 cats per 1000 people. This value is an order of magnitude lower than the estimate used by Blancher (2013). Accordingly, in the absence of any other data, the possible range of unowned cats in Canada should probably include a smaller lower bound of 150,000 (37 million humans*4/1000). While this seems very low, it is worth pointing out that only 33,491 (29%) of cats received by shelter organizations in 2016 did not have identification (HumaneCanada 2017), and could therefore possibly be previously unowned. Although, some cats in trap-neuter-release programs will also have a tattoo or chip identification.
Outdoor cats
Outdoor cats comprise two populations: owned cats that spend time outdoors, and unowned cats. We assume that all unowned cats spend all of their time outside. For owned cats, national survey data suggests that only ~30% of cats have outdoor access; however, publications concerned with conservation implications of cat predation tend to use higher values.
Loss et al. (2013) use a uniform distribution with bounds at 40 and 70% to describe the percent of owned cats with outdoor access. They claim that this value is centered on the distribution for the three nation-wide studies they cite. Of the three citations, however, two refer to the same survey (Loss et al. 2018). Of the two independent data sources, both date from the late 1990s even though there has been a consistent upward trend in the number of cats kept strictly indoors. One of the citations is for APPA survey data from 1996/7 even though more recent the APPA surveys indicate that the number of pet cats outdoors has declined steadily over the past decade (Fig 3 Rowan et al. 2019), with less than 10% outside at night, and about 30% allowed outside during the day.
Other studies support a lower percentage than used by these authors as well, especially if one considers the difference between daytime, nighttime and unsupervised access. One study of 256 US households surveyed from 1993-2003 found that only 17% of owned cats had unrestricted outdoor access, and 50% were kept indoors all the time (cited in Bernstein 2007). Similarly, in a survey of 184 cat owners at a vet hospital Clancy (2003) found 97.1% were kept in at night. Hall et al. (2016) in international survey found that US cat owners had one of the higher rates of cat confinement with only 44% allowed outdoors. Wolf (2016) in an examination of 18 surveys suggests the percent of owned cats with outdoor access is closer to 20-50%. In Canada, Blancher (2013) used a variety of research to estimate that 40 to 70% of Canadian house cats are allowed to free-roam outdoors. However, the most recent Kynetec survey (CAHI 2019) gives the percentage of owned cats permitted outside without supervision as 28%.
There are significant differences in the time of day access to the outdoors, which given hunting patterns, has implications for predation rates. Survey data also suggest that about half of these cats are allowed outdoors for no more than two to four hours each day (Clancy et al. 2003; Kays and DeWan 2004; Lord 2008). Wolf (2016) notes that is important because hunting behaviour is not continuous. Lloyd et al. (2013b) found that only 13% of 24 pet cats exhibited behaviors indicative of hunting within their first five hours outdoors. In a back-of-the-envelope calculation, Rowan (2010) suggested that cats with semi-restricted access generates about 12-15% “full time cat equivalents” of the actual hunting cat population.
We used a percent of owned cats with outdoor access of 25–50%, which encompasses the lower bound set by recent national surveys, and an upper bound as recommended by Wolf (2016). When used with the Loss et al. (2013) model, this range of owned cats with outdoor access yields a median number of annual bird kills that is ~330 million lower than the value reported by those authors.
Diet and number of prey killed
Diet
Cat predation will vary seasonally, from one geographic area and landscape type to another, and ultimately from one cat to another (Barratt 1998). That said, beside the household food, the main diet of owned cats contains small mammals (mainly rodents), small birds or insects. The main prey are generally insects and small mammals, but birds or European rabbit (e.g., Liberg 1984) are also important. Consumption of reptiles can also be important where these species are common (e.g., Loyd et al. 2013b).
Like any predator, we expect cat predation to vary with prey density (e.g., Holling 1959), increasing as prey density increases until a saturation density is reached. In particular, cats are known to be highly opportunistic (e.g., Liberg 1984), so that their predation rates will reflect both prey abundance and vulnerability. For example, Lieberg reports that during a harsh winter in Sweden cats took a very large number of rabbits that he described as dying from starvation. On islands, cats may take a large number of ground-nesting seabirds because these species are both vulnerable and abundant. Moreover, since predation rates are also related to prey density, data from over 50 years ago (i.e., before dramatic reductions in wild animal populations, and incidentally, before the wide-spread use of commercial cat food), are likely to overestimate current predation rates for owned cats.
Vulnerability will be greater where the local fauna do not share an evolutionary history with cats. In fact, Ross et al. (2019) have begun a program to familiarize native fauna with cats as predators in an attempt to mitigate the large impact they may have on Australian prey species. Therefore, predation rates on islands, or in areas such as Australia are quite unlikely to be representative of predation rates in North America. In North America, there are several native small felids, and also similar-sized native mesopredators.
Numbers
Numbers of prey items for owned cats and unowned cats are extrapolated using various methods such as examination of stomach contents, analysis of scats or owner reports. Owner reported returns are the most common method for owned cats. However, these estimates can vary quite widely, perhaps due to methods (e.g., surveys based on owner recollection vs examination of returned prey items, geographical location, cat demography, etc.). In an often cited paper, Crooks & Soulé (1999) estimate the number of cats penetrating natural habitat fragments immediately adjacent to housing developments in San Diego and their kill numbers based on responses of owner recollection. The authors indicate that cat owners reported that each outdoor cat that hunted returned on average 24 rodents, 15 birds and 17 lizards to the residence each year (or 56 prey items per cat per year). However, this figure is decidedly higher than similar studies in different locations. An examination of prey returns by 70 cats in an English village over one year yielded a total estimate of 14 prey items per cat per year (Churcher & Lawton 1987). The Mammal Society in the United Kingdom reported prey killed or captured by 964 owned cats during a five-month period in 1997 (Woods et al. 2003). The report documented more than 14,000 prey collected by cat owners from their animals, and also yielded a mean estimate of 14 items per owned cat per year. A study in Cape Town (Morling 2014) also gives prey returns of 14 items per year. In a study in Switzerland, Tschanz and coauthors (2011) note that a large fraction of owners considerably overestimated their cat’s predation, indicating that surveying predation rates by means of a retrospective questionnaire alone is not sufficient, and possibly explaining the discrepancy between the Crooks & Soule (1999) estimate and other studies that relied on a closer reporting of prey return items.
Owned cat predation rates
There are two major studies on the topic of owned cat predation in North America. Blancher (2013) estimated that owned cats returned 2.8 to 14 birds per year range, with a higher value for rural cats than urban cats. Loss et al. (2013) provide a range of 4.2 to 18.3 birds per cat per year based on US and European region studies. (Note that both authors adjust these numbers upwards, for predation events not detected by owners). Loss et al. (2013) provide estimated numbers for rodents as well. For mammal kills by owned cats, they give 8.7 to 21.8 items (Loss et al. 2013).
Loss et al. (2013, suppl) and Blancher (2013) provide the references used to inform the choices they made for the range and distribution of predation rates of owned and unowned cats. We used these lists as a starting point for our dataset of owned cat predation rates, excluding all references that had been excluded for various reasons by either of the two sets of authors (e.g., small sample size, survey data that required recollection of past events). Because cats are opportunistic predators (e.g., Liberg 1984), like these authors, we excluded data from islands, where food choices would have been limited compared to mainland locations. We also excluded data from Australia and New Zealand, where prey may not have an evolutionary history that included small predators like cats. In general, prey without this history are less able to escape predation, although there are promising exposure techniques to increase prey survival rates (e.g., Ross et al. 2019).
We note in passing that that there is a significant negative relationship between year of data collection and reported predation rates. We remind the reader that predation rates are determined by prey density, that wild bird and mammal populations have declined dramatically in the past century, and commercial cat food was not in common use until some years after World War II. While we did not exclude data based on year of collection, it seems unlikely that current predation rates for owned cats are well described by data prior to 1950.
The raw data from these papers used by Loss et al. (2013) or Blancher (2013) is not given in either publication. Instead, these authors both give calculated annual rates based on corrections for seasonal variation in predation success. Loss et al. (2013) state that the adjusted partial-year predation estimates are calculated using the average proportion of prey taken in each month from 4 different studies of bird predation, 2 of which include data for mammals (Churcher & Lawton 1987; Barratt 1997; Fiore & Sullivan unpub; van Heezik et al. 2010). These proportions are not reported. Blancher (2013) used the same approach for birds and a similar selection of year round studies. However, this author does report the proportions used in a figure (Fig 1 Blancher (2013)). As a result, it can be difficult to determine what data from a given study Loss et al. (2013) or Blancher (2013) actually used in their reporting. We provide some commentary below on references that we decided to exclude, but which were used by these authors in their calculations.
We used a combination of the papers from Loss et al. (2013) and Blancher (2013) that we did not exclude, and a few newer papers that we uncovered to create a set of 7 estimates of owned cat predation rates. We partitioned these rates between bird and mammal kills to allow comparison to the literature. The calculated mean annual predation rates, unadjusted for season of data collection, were fit to lognormal, normal and uniform distributions to determine the best parameters for simulations. For birds, a lognormal distribution was a good descriptor, but for mammals there were no clear best fits (Fig 4), so we selected a lognormal distribution for each group, because of the advantage over the normal distribution of excluding negative values, and the advantage over the uniform distribution of not suggesting even probability for all values (which does not seem to be true for birds).
Predation studies used for owned cat predation data in Loss et al. (2013) or Blancher (2013) that we excluded
As previously mentioned, we excluded all data that had been excluded by either Loss et al. (2013) or Blancher (2013). We also excluded data from Australia, New Zealand and islands. Below we give our reasons for excluding several papers included by these two sets of authors that do not fall under these criteria.
McMurray and Sperry 1941: The largest reported predation rate for owned cats in Loss et al. (2013) at 33.18 birds/year is derived from this paper on the basis of 2 birds found in the stomachs of 22 cats, some of which may have been pets. However, Blancher (2013) used this paper as a data point for unowned cats only at 48 birds per year. Our value is 31 birds per unowned cat per year.
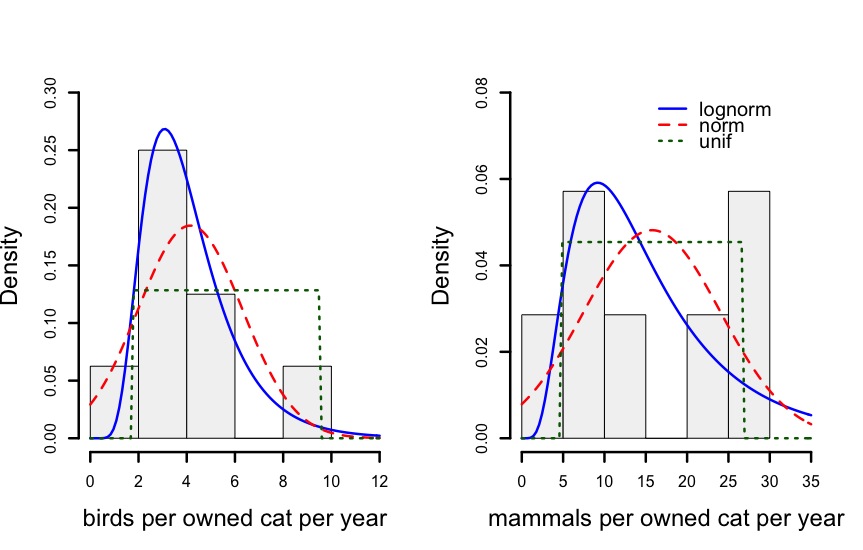
Figure 4: Histograms of annual estimated predation rates from literature data, and lognormal, normal and uniform distributions to this data via maximum likelihood. (Please see https://github.com/kcudding/ReThinkCats/ for references and raw data.)
The paper deals with stomach contents from three different groups: Wichita Mountains Wildlife Refuge (n=24), the Fort Sill Military Reservation (22 from residential sections and 13 from non- residential areas), and those from other locations in Oklahoma (25). Twelve of the 107 stomachs were empty and 11 more contained only traces of food, so percentage tabulations were based on the analyses of 84 stomachs.
The three sites are described as:
Wichita Mountains Wildlife Refuge (24 samples): Sampling dates not indicated. Includes unowned cats as well as some animals described as household pets abandoned at camp grounds. Birds reported as “trace”. Mammals as 54% weight.
Fort Sill Military Reservation (22 +13): There are two sampling locations here: “residential” and “non-residential” sections. Authors state that most cats from residential sections were trapped during the fall of 1939. Trapping in both sites described as fall and winter from mid-Oct to mid-Jan. Authors state that cats from residential sections “probably include many so-called ‘alley cats’ as well as a few house pets”. Two birds, an English sparrow and a meadowlark, formed the bulk of the last meal of 2 cats from residential sections and made an average of 6.5 per cent of the food for the lot (so they are reporting % mass). Only 1 of the 13 cats from nonresidential areas captured a bird, a robin.
Cats Taken in Other Sections of Oklahoma (25 stomachs): Other samples came from 12 counties collected throughout the year and probably include some domestic pets that were hunting along roadsides at night as well as more unowned individuals. Birds composed 6 per cent of the food and occurred in 5 of the 25 stomachs. Remains of 3 birds – a Brewer's blackbird, a horned lark, and a meadowlark – were found in one stomach collected in February.
As far as we can tell, Loss et al. (2013) used the data of 2 birds in 22 stomachs from the Fort Sill Military reservation as “Owned”. These cats are described by the authors as including “a few house pets” and “alley cats”. The unowned data Loss and coauthors report is then the sum of Wichita Mountains Wildlife Refuge cats, and the cats trapped in the nonresidential areas of Fort Sill Military Reservation. Since the authors of the original paper do not know what percentage of their cat samples that were owned cats, and report simply that the one sample “probably included a few house pets”, this does not seem a good classification. Moreover, only including full stomachs will overestimate predation success. We record data from this paper as 9 birds in 107 stomachs, to give an overall predation rate of 0.084 birds/day for (31 per year) for primarily unowned cats in rural areas in 1941. Therefore, we do not use this data as an estimate for owned cat predation. We note that it is not possible to get predation rates for mammals from this paper given that only percent mass is reported, not percent occurrence.
Eberhard (1954): This author examined 202 cat stomachs and reported on 121 stomachs that contained prey items. Cats were taken by gathering of motor traffic kills, shooting, and trapping from 22 counties throughout the state, the majority having been collected from Centre, Montgomery, and Bucks Counties of Pennsylvania. Most specimens came from farm areas, although a lesser number was taken from small towns. Those cats which were recovered or captured in fields were tagged "field," and those animals which were thought to be pets or were taken in towns were tagged "non-field”. Data were recorded separately for spring and summer, vs. fall and winter.
Loss et al. (2013) have apparently classified the “non-field” cats as owned and used the frequency of bird remains occurrence reported by Eberhard (birds found in 4 out of 47 “non-field” cats stomachs, and field cats as unowned birds in 24 out of 107 stomachs). Cats included in the owned category almost certainly contained many unowned cats since unowned cat population density is highly correlated with human population density. In addition, 48 stomachs were excluded from the study because they were empty. If this study is to be used is would probably be better to report as 28 stomachs out of 202 bird remains (assumed predation rate of 0.14 birds/day or 51 birds per year) for rural areas (farms and small towns) as probably unowned cats in 1949-1951.
Loss et al. (2013) describe their calculated rate of 124 rodents per year (found in 16 out of 47 cat stomachs) for non-field cats as abnormally high, and exclude this data for owned cats, but retain the value of 297 per year for unowned cats. Given the classification information and the excluded stomachs, it seems better to report this data as mammal remains found in 103 out of 202 stomachs (predation rate of 0.51 mammals/day or 186 mammals per year per cat) as probably unowned cats in rural areas from 1949 to 1951.
Liberg (1984): The author reports the frequency of occurrence of bird or mammal remains in scat collected from 1974 to 1979 in a rural Swedish village. Scat is attributed to unowned or owned cats on the basis of location of collection and in some cases visual id. The author reports that about half the bird biomass in scats was Galliformes (e.g., chickens and pheasants). As a result we were unclear as to how to distinguish kills of domesticated farm chickens from kills of wild birds, and we have therefore excluded the bird data from this paper. Loss et al. (2013) do not include the mammal data from this paper, and our calculations suggest that the mammal predation rate would fall into the category of “abnormally high” which Loss and coauthors used as grounds for excluding several other studies. Therefore, we have excluded the mammal data as well.
Nelson et al. 2005: The paper reports on prey returns for cats known to be hunters wearing either regular collars or collars fitted with various sound devices to decrease predation. Volunteers were recruited from across the UK (mainly England) through advertisements in various cat and wildlife magazines, and a website. Cat owners were selected to take part in the project if they reported that their cat(s) killed at least one prey item a week and did not currently wear a collar.
Of the original sample of 150 original volunteers, 16 dropped out of the study, while 62 failed to return any prey. Since the study aimed at evaluating the efficacy of deterrent devices on active hunters, and since the authors could not distinguish between failure to complete the survey and predation failure, they excluded these 62 samples. As a result, they also conclude: “The study was not designed using a random sample of cats; indeed cats were selected for their tendency to hunt. In view of this, it would be completely inappropriate to estimate national kill rates by extrapolating from the data reported here.” Therefore, we have excluded this data.
Coleman and Temple (1996): This reference is not a peer-reviewed scientific publication and cites unknown data (i.e., there is no citation list or even names and dates for data cited as “four other studies”). Further, there are no predation data nor methods described. Other authors have critiqued the use of this reference, and have concluded that the other other data referred to may be from Errington (1936), Parmalee (1953), and Eberhard (1954). This conclusion was possibly made by referring to another paper: Coleman, J. S., & Temple, S. A. (1995). How many birds do cats kill. Wildlife Control Technology, 44, which we cannot locate. Wolf (2016) claims that the three mentioned studies were already included in the Loss et al. (2013) data set, and that the fourth (Mitchell and Beck 1992) was excluded by Loss et al. (2013) due to its small sample size. The 4-year predation study for Wisconsin that the authors mention may be either a survey on cat ownership they conducted, or some unpublished study. Loss and coauthors (2013) use this paper for unowned cat predation rates while Blancher (2013) uses it for rural owned cats. However, as far as we can tell, this paper does not include original data and has been excluded.
Baker et al. (2005): Loss et al. (2013) use both this paper and Baker et al. (2008), however, the 2008 paper, in part, reports on the 2005 data. As a result, we have used only the 2005 data reported in Baker et al. (2008). Blancher (2013) only reports on the 2008 paper, and gives a comparatively low bird predation rate (0.6 birds/year, whereas we calculate 1.75 birds per year without seasonal adjustment for both the Baker (2005) and the Baker (2008) studies). We have excluded this paper because of possible duplication.
Percentage of outdoor owned cats that hunt
In all of the studies of owned cat predation, there are some number for which predation is not detected. This may be because cats do not necessarily return prey to owners, owners do not respond to requests for information, the sampling interval is too short to detect infrequent predation events, or because the cats do not successfully hunt. For example, Baker et al. (2008) report that only 40% of cats in the study returned any prey to their owners; however, this absence does not necessarily indicate failure to hunt. Loyd and coauthors (2013b) placed video cameras on 55 cats and obtained on average of 7-10 days of total footage. Of the 55 cats, only 24 were observed to engage in “predation behaviour” of stalking or chasing prey, and only 16 (~30%) were observed to actually capture prey.
In addition, studies have indicated that there is a lognormal distribution of observed predation success among individual cats, with a few individuals acting as “super-predators”, and a much larger number capturing very few or no prey. Lloyd et al. (2013b) report that of their group of 55 cats, one individual captured five prey, and 3 captured 4, while the majority captured one to no prey. Similarly, in a study in Switzerland, five cats accounted for 75% of prey and 11 cats did not return any prey items (Tschanz et al. 2011).
Loss et al. (2013) cite three references for their use of a uniform distribution between 0.5 and 0.8 to describe the percentage of owned cats that hunt (Barratt et al. 1998, Fiore unpub, Crooks & Soule 1999), while Blancher (2013) assumes that all outdoor cats hunt successfully. This factor may be a great importance when we consider that, by using the mean predation rate to summarize data in the published studies, we may have overestimated the average predation rate per cat (because of the influence of the long tail of the distribution on the mean). Barratt et al. (1998) note that using the mean instead of the median will double the estimated per cat predation rate. Further, when constructing simulations Loss et al. (2013) and Blancher (2013) use a uniform distribution to describe possible predation rates, which assumes that both large and small rates are equally probable: an unlikely scenario given what we know about predation rates.
Prey return rate
The prey return rate is a correction factor designed to account for the fact that predation rates estimated from the number of prey returned to the owners only represents a portion of the total kills. Next to the number of unowned cats, this is the model parameter for which we have the least information. Moreover, a closer examination of the data indicates that the use of this parameter will generate maximum owned cat predation rates greater than the maximum unowned cat predation rates in the original Loss et al. model (2013) based on EU and US data, and the same is true for the rural cat predation rate in the Blancher (2013) model, possibly because in both cases the correction factor is used across a range selected from data that included scats and stomach contents as well as prey returns.
Loss et al. (2013) use a uniform distribution between 1.2 to 3.3 to estimate of this parameter, based on three studies. Wolf (2016) notes that the lowest estimate (from George 1974) is actually based on a misreading. The author of that paper is accounting for lack of continuous observation of cat return rates by the human owner rather than representing a difference between prey captures and prey returns. The top of the range used by Loss et al. (2013) is based on a predation rate calculated from 4 successful mammal captures noted in 181 total hours of observations of 12 cats in New York (Kays & Dewan 2004), and comparing the extrapolated predation rate to prey returns. Blancher (2013) also use a misreading for the George (1974) study for the bottom of a uniform distribution ranging from 1.2 to 5.8 for this parameter. They note the unsatisfactory sample size from Kays & Dewan (2004), and report on an unpublished manuscript by Fiore and Sullivan, where the authors analyzed scat samples to show that 21% of the time, cats ingested birds without owner knowledge, data from Loyd et al. (2013b) from video cameras on cats (n = 39) and found that pet cats brought home 23% of prey. Krauze-Gryz et al. (2012) found that predation rate for all prey types was 11.4 times higher when based on scats and stomach samples than from prey returns (including insect consumption). While the published paper reports no significant difference between prey returns and consumption for birds, Blancher (2013) states “their raw data for birds suggest a 5.8x adjustment”, and use this value as the maximum of a uniform distribution from 1.2 to 5.8 for both birds.
References
AAPOR Report. Prepared for the AAPOR Executive Council by a Task Force operating under the auspices of the AAPOR Standards Committee, with members including:, Baker, R., Blumberg, S. J., Brick, J. M., Couper, M. P., Courtright, M., ... & Groves, R. M. (2010). Research synthesis: AAPOR report on online panels. Public Opinion Quarterly, 74(4), 711-781. https://pprg.stanford.edu/wp-content/uploads/2010-AAPOR-Report-on-Online-Panels.pdf
Alhadeff, A. L., Su, Z., Hernandez, E., Klima, M. L., Phillips, S. Z., Holland, R. A., ... & Betley, J. N. (2018). A neural circuit for the suppression of pain by a competing need state. Cell, 173(1), 140-152. https://doi.org/10.1016/j.cell.2018.02.057
Amat, M., de la Torre, J. L. R., Fatjó, J., Mariotti, V. M., Van Wijk, S., & Manteca, X. (2009). Potential risk factors associated with feline behaviour problems. Applied Animal Behaviour Science, 121(2), 134-139. https://doi.org/10.1016/j.applanim.2009.09.012
Bain, M., & Stelow, E. (2014). Feline aggression toward family members: A guide for practitioners. Veterinary Clinics of North America - Small Animal Practice, 44(3), 581-597. https://doi.org/10.1016/j.cvsm.2014.01.001
Baker, P. J., Bentley, A. J., Ansell, R. J., & Harris, S. (2005). Impact of predation by domestic cats Felis catus in an urban area. Mammal Review, 35(3‐4), 302-312. https://doi.org/10.1111/j.1365-2907.2005.00071.x
Baker, P. J., Molony, S. E., Stone, E., Cuthill, I. C., & Harris, S. (2008). Cats about town: is predation by free‐ranging pet cats Felis catus likely to affect urban bird populations?. Ibis, 150, 86-99. https://doi.org/10.1111/j.1474-919X.2008.00836.x
Barratt, D. G. (1997). Predation by house cats, Felis catus (L.), in Canberra, Australia. I. Prey composition and preference. Wildlife Research, 24(3), 263-277. https://doi.org/10.1071/WR96020
Barratt, D. G. (1998). Predation by house cats, Felis catus (L.), in Canberra, Australia. II. Factors affecting the amount of prey caught and estimates of the impact on wildlife. Wildlife Research, 25(5), 475-487. https://doi.org/10.1071/WR97026
Beckerman, A. P., Boots, M., & Gaston, K. J. (2007). Urban bird declines and the fear of cats. Animal Conservation, 10(3), 320-325. https://doi.org/10.1111/j.1469-1795.2007.00115.x
Bernstein, P. L. (2007). The human-cat relationship. In The welfare of cats I. Rochlitz (Ed.), pp. 47-89. Springer, Dordrecht. https://www.gwern.net/docs/catnip/2005-bernstein.pdf
Berry, R. J. (1981). Town mouse, country mouse: adaptation and adaptability in Mus domesticus (M. musculus domesticus). Mammal Review, 11(3), 91-136. https://doi.org/10.1111/j.1365-2907.1981.tb00001.x
Berry, R. J., & Jakobson, M. E. (1971). Life and death in an island population of the house mouse. Experimental Gerontology, 6(2), 187-197. https://doi.org/10.1016/S0531-5565(71)80018-9
Berry, R. J., Jakobson, M. E., & Triggs, G. S. (1973). Survival in wild‐living mice. Mammal Review, 3(2), 46-57. https://onlinelibrary.wiley.com/doi/pdf/10.1111/j.1365-2907.1973.tb00171.x
Biben, M. (1979). Predation and predatory play behaviour of domestic cats. Animal Behaviour, 27, 81-94. https://doi.org/10.1016/0003-3472(79)90129-5
Blancher, P. (2013). Estimated number of birds killed by house cats (Felis catus) in Canada. Avian Conservation and Ecology, 8(2). http://www.ace-eco.org/vol8/iss2/art3/
Buffington, C. T. (2002). External and internal influences on disease risk in cats. Journal of the American Veterinary Medical Association, 220(7), 994-1002. https://doi.org/10.2460/javma.2002.220.994
CAHI (2019). Latest Canadian Pet Population Figures Released. https://www.cahi-icsa.ca/press-releases/latest-canadian-pet-population-figures-released
Chalkowski, K., Wilson, A. E., Lepczyk, C. A., & Zohdy, S. (2019). Who let the cats out? A global meta-analysis on risk of parasitic infection in indoor versus outdoor domestic cats (Felis catus). Biology Letters, 15(4), 20180840. https://doi.org/10.1098/rsbl.2018.0840
Chamberlain, D. E., Cannon, A. R., Toms, M. P., Leech, D. I., Hatchwell, B. J., & Gaston, K. J. (2009). Avian productivity in urban landscapes: a review and meta‐analysis. Ibis, 151(1), 1-18. https://doi.org/10.1111/j.1474-919X.2008.00899.x
Churcher, P. B., & Lawton, J. H. (1987). Predation by domestic cats in an English village. Journal of Zoology, 212(3), 439-455. https://doi.org/10.1111/j.1469-7998.1987.tb02915.x
Clancy, E. A., Moore, A. S., & Bertone, E. R. (2003). Evaluation of cat and owner characteristics and their relationships to outdoor access of owned cats. Journal of the American Veterinary Medical Association, 222(11), 1541-1545. https://doi.org/10.2460/javma.2003.222.1541
Coleman, J. S., & Temple, S. A. (1996). On the prowl. Wisconsin Natural Resources, 20(6), 4-8. https://dnr.wi.gov/wnrmag/html/stories/1996/dec96/cats.htm
Crooks, K. R., & Soulé, M. E. (1999). Mesopredator release and avifaunal extinctions in a fragmented system. Nature, 400(6744), 563. https://www.nature.com/articles/23028
Crowley, S. L., Cecchetti, M., & McDonald, R. A. (2019). Hunting behaviour in domestic cats: An exploratory study of risk and responsibility among cat owners. People and Nature, 1(1), 18-30. https://doi.org/10.1002/pan3.6
Cypher, B. L., McMillin, S. C., Westall, T. L., Van Horn Job, C., Hosea, R. C., Finlayson, B. J., & Kelly, E. C. (2014). Rodenticide exposure among endangered kit foxes relative to habitat use in an urban landscape. Cities and the Environment (CATE), 7(1), 8. http://digitalcommons.lmu.edu/cate/vol7/iss1/8
Dauphiné, N. I. C. O., & Cooper, R. J. (2009, October). Impacts of free-ranging domestic cats (Felis catus) on birds in the United States: a review of recent research with conservation and management recommendations. In Proceedings of the fourth international partners in flight conference: tundra to tropics (Vol. 205). http://www.birdsphotography.com/cats/by_dauphine_cooper.pdf
Dickman, C. & Denny E. (2010). Strategies to reduce conflict: managing feral and stray cats. In: Proceedings of the RSPCA Scientific Seminar. Australia: RSPCA: 41–45. Tensen, M. & Jones, B. (Eds). https://www.researchgate.net/publication/330038825_Strategies_to_reduce_conflict_managing_feral_and_stray_cats/link/5c2b183fa6fdccfc707509a1/download
Dierschke, V. (2003). Predation hazard during migratory stopover: are light or heavy birds under risk?. Journal of Avian Biology, 34(1), 24-29.https://doi.org/10.1034/j.1600-048X.2003.03049.x
Eberhard, T. (1954). Food habits of Pennsylvania house cats. The Journal of Wildlife Management, 18(2), 284-286. https://www.jstor.org/stable/3797736
Erickson, W. A., & Urban, D. J. (2004). Potential risks of nine rodenticides to birds and nontarget mammals: a comparative approach (p. 225). Washington, DC: US Environmental Protection Agency, Office of Prevention, Pesticides and Toxic Substances. http://pesticideresearch.com/site/docs/bulletins/EPAComparisonRodenticideRisks.pdf
Errington, P. L. (1936). Notes on food habits of southern Wisconsin house cats. Journal of Mammalogy, 17(1), 64-65. https://doi.org/10.1093/jmammal/17.1.64-b
Food and Agriculture Organization of the United Nations (FAO). (2019). FAOSTAT Database. Rome, Italy: FAO. Retrieved from http://www.fao.org/faostat/en/#home
Fitzgerald, B.M. & Turner, C.T. (2000). Hunting behaviour of domestic cats and their impact on prey populations. In: The domestic cat. The biology of its behaviour. 2nd edn: 151– 176. Turner, D.C. & Bateson, P. (Eds). Cambridge: Cambridge University Press. https://www.gwern.net/docs/catnip/2000-turner-thedomesticcat.pdf
Fiore, C. A., & Sullivan, K. B. (2000). Domestic cat (Felis catus) predation of birds in an urban environment. Wichita, KS: Wichita State University.
Flockhart, D. T. T., Norris, D. R., & Coe, J. B. (2016). Predicting free‐roaming cat population densities in urban areas. Animal Conservation, 19(5), 472-483. https://doi.org/10.1111/acv.12264
Flux, J. E. (1993). Relative effect of cats, myxomatosis, traditional control, or competitors in removing rabbits from islands. New Zealand Journal of Zoology, 20(1), 13-18. https://doi.org/10.1080/03014223.1993.10423238
Gabriel, M., Diller, L., Dumbacher, J., Wengert, G., Higley, J., Poppenga, R., & Mendia, S. (2018). Exposure to rodenticides in Northern Spotted and Barred Owls on remote forest lands in northwestern California: evidence of food web contamination. Avian Conservation and Ecology, 13(1). https://doi.org/10.5751/ACE-01134-130102
Genovesi, P., Besa, M., & Toso, S. (1995). Ecology of a feral cat Felis catus population in an agricultural area of northern Italy. Wildlife biology, 1(1), 233-238. https://doi.org/10.2981/wlb.1995.0028
Gibson, E., Blackwell, E., Roberts, C., Gruffydd-Jones, T., Murray, J., & Williams, J. (2018). Comparison of diseases and problematic behaviours in cats confined indoors or allowed outdoor access. In: BSAVA Congress Proceedings 2018 (pp. 471-471). BSAVA Library. https://www.bsavalibrary.com/docserver/fulltext/10.22233/9781910443590/9781910443590.73.3.pdf
Glass, G. E., Gardner-Santana, L. C., Holt, R. D., Chen, J., Shields, T. M., Roy, M., ... & Klein, S. L. (2009). Trophic garnishes: Cat–rat interactions in an urban environment. PLoS One, 4(6), e5794. https://doi.org/10.1371/journal.pone.0005794
Gurevitch, J., Morrow, L. L., Wallace, A., & Walsh, J. S. (1992). A meta-analysis of competition in field experiments. The American Naturalist, 140(4), 539-572. https://doi.org/10.1086/285428
Hansen, C. M., Paterson, A. M., Ross, J. G., & Ogilvie, S. C. (2018). Estimating feral cat (Felis catus) density in a rural to urban gradient using camera trapping. New Zealand Journal of Zoology, 45(3), 213-226. https://doi.org/10.1080/03014223.2018.1494609
Hall, C. M., Fontaine, J. B., Bryant, K. A., & Calver, M. C. (2015). Assessing the effectiveness of the Birdsbesafe® anti-predation collar cover in reducing predation on wildlife by pet cats in Western Australia. Applied Animal Behaviour Science, 173, 40-51. https://doi.org/10.1016/j.applanim.2015.01.004
Hall, C. M., Adams, N. A., Bradley, J. S., Bryant, K. A., Davis, A. A., Dickman, C. R., ... & Pollock, K. H. (2016). Community attitudes and practices of urban residents regarding predation by pet cats on wildlife: an international comparison. PloS one, 11(4), e0151962. https://doi.org/10.1371/journal.pone.0151962
Hand, A. (2019). Estimating feral cat densities using distance sampling in an urban environment. Ecology and Evolution, 9(5), 2699-2705. https://onlinelibrary.wiley.com/doi/pdf/10.1002/ece3.4938
Heske, E. J., Brown, J. H., & Mistry, S. (1994). Long‐term experimental study of a Chihuahuan Desert rodent community: 13 years of competition. Ecology, 75(2), 438-445. https://doi.org/10.2307/1939547
Hindmarch, S., Elliott, J. E., & Morzillo, A. (2018). Rats! What triggers us to control for rodents? Rodenticide user survey in British Columbia, Canada. International Journal of Environmental Studies, 75(6), 1011-1030. https://www.tandfonline.com/doi/citedby/10.1080/00207233.2018.1479565
Holling, C. S. (1959). Some characteristics of simple types of predation and parasitism. The Canadian Entomologist, 91(7), 385-398. https://doi.org/10.4039/Ent91385-7
HumaneCanada (2017). Cats in Canada 2017: A Five-Year Review of Cat Overpopulation. https://www.humanecanada.ca/cats_in_canada_2017
Karl, B.J. and Best, H.A. (1982) Feral cats on Stewart Island; their foods, and their effects on
kakapo. N. Z. J. Zool. 9, 287–294 https://www.tandfonline.com/doi/pdf/10.1080/03014223.1982.10423857
Kays, R. W., & DeWan, A. A. (2004). Ecological impact of inside/outside house cats around a suburban nature preserve. In Animal Conservation forum (Vol. 7, No. 3, pp. 273-283). Cambridge University Press. https://doi.org/10.1017/S1367943004001489
Krauze‐Gryz, D., Gryz, J., & Goszczyński, J. (2012). Predation by domestic cats in rural areas of central P oland: an assessment based on two methods. Journal of Zoology, 288(4), 260-266. https://doi.org/10.1111/j.1469-7998.2012.00950.x
Laurie, E. M. O. (1946). The reproduction of the house-mouse (Mus musculus) living in different environments. Proceedings of the Royal Society of London. Series B-Biological Sciences, 133(872), 248-281. https://doi.org/10.1098/rspb.1946.0012
Legge, S., Murphy, B. P., McGregor, H., Woinarski, J. C. Z., Augusteyn, J., Ballard, G., ... & Edwards, G. (2017). Enumerating a continental-scale threat: how many feral cats are in Australia?. Biological Conservation, 206, 293-303. https://doi.org/10.1016/j.biocon.2016.11.032
Lepczyk, C. A., Mertig, A. G., & Liu, J. (2004). Landowners and cat predation across rural-to-urban landscapes. Biological conservation, 115(2), 191-201. https://doi.org/10.1016/S0006-3207(03)00107-1
Levy, J. K., Woods, J. E., Turick, S. L., & Etheridge, D. L. (2003). Number of unowned free-roaming cats in a college community in the southern United States and characteristics of community residents who feed them. Journal of the American Veterinary Medical Association, 223(2), 202-205. https://doi.org/10.2460/javma.2003.223.202
Liberg, O. (1984). Food habits and prey impact by feral and house-based domestic cats in a rural area in southern Sweden. Journal of Mammalogy, 65(3), 424-432. https://doi.org/10.2307/1381089
Liberg, O., Sandell, M., Pontier, D., & Natoli, E. (2000). Density, spatial organisation and reproductive tactics in the domestic cat and other felids. The domestic cat: the biology of its behaviour. Cambridge University Press, Cambridge, United Kingdom, 119-147. https://books.google.ca/books?hl=en&lr=&id=GgUwg6gU7n4C
Lord, L. K., Griffin, B., Slater, M. R., & Levy, J. K. (2010). Evaluation of collars and microchips for visual and permanent identification of pet cats. Journal of the American Veterinary Medical Association, 237(4), 387-394. https://doi.org/10.2460/javma.237.4.387
Loss, S. R., Will, T., & Marra, P. P. (2013). The impact of free-ranging domestic cats on wildlife of the United States. Nature communications, 4, 1396. https://www.nature.com/articles/ncomms2380
Loss, S. R., Will, T., & Marra, P. P. (2015). Direct mortality of birds from anthropogenic causes. Annual Review of Ecology, Evolution, and Systematics, 46, 99-120. https://repository.si.edu/bitstream/handle/10088/27854/NZP_Marra_2015-Direct_Mortality_of_Birds_from_Anthropogenic_Causes.pdf
Loss, S. R., Will, T., Longcore, T., & Marra, P. P. (2018). Responding to misinformation and criticisms regarding United States cat predation estimates. Biological invasions, 20(12), 3385-3396. https://link.springer.com/article/10.1007/s10530-018-1796-y
Lord, L. K. (2008). Attitudes toward and perceptions of free-roaming cats among individuals living in Ohio. Journal of the American Veterinary Medical Association, 232(8), 1159-1167.https://doi.org/10.2460/javma.232.8.1159
Loyd, K. A. T., Hernandez, S. M., Abernathy, K. J., Shock, B. C., & Marshall, G. J. (2013a). Risk behaviours exhibited by free-roaming cats in a suburban US town. Veterinary Record, 173, 29. http://dx.doi.org/10.1136/vr.101222
Loyd, K. A. T., Hernandez, S. M., Carroll, J. P., Abernathy, K. J., & Marshall, G. J. (2013b). Quantifying free-roaming domestic cat predation using animal-borne video cameras. Biological Conservation, 160, 183-189. https://doi.org/10.1016/j.biocon.2013.01.008
Lynn, W. (2015). Australia’s war on feral cats: shaky science, missing ethics. https://theconversation.com/australias-war-on-feral-cats-shaky-science-missing-ethics-47444.
Lynn, W. S., Santiago‐Ávila, F., Lindenmayer, J., Hadidian, J., Wallach, A., & King, B. J. (2019). A moral panic over cats. Conservation Biology 33: 769-776. https://doi.org/10.1111/cobi.13346
Malpass, J. S., Rodewald, A. D., Matthews, S. N., & Kearns, L. J. (2018). Nest predators, but not nest survival, differ between adjacent urban habitats. Urban Ecosystems, 21(3), 551-564.https://doi.org/10.1007/s11252-017-0725-7
Mason, G. J. & K. E. Littin (2003). The humaneness of rodent pest control. Animal Welfare 12, 1 -37. https://www.researchgate.net/profile/Kate_Littin/publication/253097632_The_humaneness_of_rodent_pest_control/links/0c96053c5ca400bd7b000000.pdf
McDonald, J. L., Cleasby, I. R., Brodbelt, D. C., Church, D. B., & O'neill, D. G. (2017). Mortality due to trauma in cats attending veterinary practices in central and south‐east England. Journal of Small Animal Practice, 58(10), 570-576. https://doi.org/10.1111/jsap.12716
McMurry, F. B., & Sperry, C. C. (1941). Food of feral house cats in Oklahoma, a progress report. Journal of Mammalogy, 22(2), 185-190. https://www.jstor.org/stable/1374915
Memmott, K., Murray, M., & Rutberg, A. (2017). Use of anticoagulant rodenticides by pest management professionals in Massachusetts, USA. Ecotoxicology, 26(1), 90-96. https://doi.org/10.1007/s10646-016-1744-5
Mitchell, J. C., & Beck, R. A. (1992). Free-ranging domestic cat predation on native vertebrates in rural and urban Virginia. Virginia Journal of Science, 43(1B), 197-207.
Møller, A. P., & Erritzøe, J. (2000). Predation against birds with low immunocompetence. Oecologia, 122(4), 500-504. https://doi.org/10.1007/s004420050972
Morling, F. (2014). Cape Town's cats: reassessing predation through kitty-cams (Doctoral dissertation, University of Cape Town). https://open.uct.ac.za/bitstream/handle/11427/9099/thesis_sci_2014_morling_f.pdf?sequence=1
Morzillo, A. T., & Mertig, A. G. (2011). Urban resident attitudes toward rodents, rodent control products, and environmental effects. Urban Ecosystems, 14(2), 243-260. https://doi.org/10.1007/s11252-010-0152-5
Morzillo, A. T., & Schwartz, M. D. (2011). Landscape characteristics affect animal control by urban residents. Ecosphere, 2(11), 1-16. https://doi.org/10.1890/ES11-00120.1
Murray, M. (2017). Anticoagulant rodenticide exposure and toxicosis in four species of birds of prey in Massachusetts, USA, 2012–2016, in relation to use of rodenticides by pest management professionals. Ecotoxicology, 26(8), 1041-1050. https://doi.org/10.1007/s10646-017-1832-1
Natoli, E. (1985). Spacing pattern in a colony of urban stray cats (Felis catus L.) in the historic centre of Rome. Applied Animal Behaviour Science, 14(3), 289-304. https://doi.org/10.1016/0168-1591(85)90009-7
Nelson, S. H., Evans, A. D., & Bradbury, R. B. (2005). The efficacy of collar-mounted devices in reducing the rate of predation of wildlife by domestic cats. Applied Animal Behaviour Science, 94(3-4), 273-285. https://doi.org/10.1016/j.applanim.2005.04.003
Nogeire, T. M., Lawler, J. J., Schumaker, N. H., Cypher, B. L., & Phillips, S. E. (2015). Land use as a driver of patterns of rodenticide exposure in modeled kit fox populations. PloS one, 10(8), e0133351. https://doi.org/10.1371/journal.pone.0133351
Öhlund, M., Egenvall, A., Fall, T., Hansson‐Hamlin, H., Röcklinsberg, H., & Holst, B. S. (2017). Environmental risk factors for diabetes mellitus in cats. Journal of Veterinary Internal Medicine, 31(1), 29-35. https://onlinelibrary.wiley.com/doi/pdf/10.1111/jvim.14618
Parmalee, P. W. (1953). Food habits of the feral house cat in east-central Texas. The Journal of Wildlife Management, 17(3), 375-376.
Piquet, J.C., Baumgartner, E.S., Medina, F.M., Díaz-Luis, N., Sevilla, J., López, H., Nogales, M. and López-Darias, M., 2019. A resource-efficient procedure to improve planning of invasive cat management on inhabited islets. Biological Invasions, 21(5), pp.1817-1831. https://doi.org/10.1007/s10530-019-01941-x
Pocock, M. J., Searle, J. B., & White, P. C. (2004). Adaptations of animals to commensal habitats: population dynamics of house mice Mus musculus domesticus on farms. Journal of Animal Ecology, 73(5), 878-888. https://besjournals.onlinelibrary.wiley.com/doi/pdf/10.1111/j.0021-8790.2004.00863.x
Regnery, J., Parrhysius, P., Schulz, R. S., Möhlenkamp, C., Buchmeier, G., Reifferscheid, G., & Brinke, M. (2019). Wastewater-borne exposure of limnic fish to anticoagulant rodenticides. Water Research, 115090. https://doi.org/10.1007/s10311-018-0788-6
Ross, A. K., Letnic, M., Blumstein, D. T., & Moseby, K. E. (2019). Reversing the effects of evolutionary prey naiveté through controlled predator exposure. Journal of Applied Ecology 56: 1761-1769. https://doi.org/10.1111/1365-2664.13406
Rowan, A. (2010). Cat demographics. Humane Society. https://www.humanesociety.org/sites/default/files/docs/2012-rowan-cat-trends-usa.pdf
Rowan, A. (2018). Companion Animal Statistics in the USA The Humane Society Institute for Science and Policy Animal Studies Repository. Companion Animal Statistics in the USA https://animalstudiesrepository.org/demscapop/7
Rowan, A. N., Kartal, T., & Hadidian, J. (2019). Cat Demographics & Impact on Wildlife in the USA, the UK, Australia and New Zealand: Facts and Values. Journal of Applied Animal Ethics Research, 1(aop), 1-31. https://doi.org/10.1163/25889567-12340013
Rowe, A. (2018) Adoption Level Advocacy: A Cost-Effective Program to Reduce Wild Animal Suffering. https://www.utility.farm/words/adoption-level-advocacy
Schoener, T. W. (1983). Field experiments on interspecific competition. The American Naturalist, 122(2), 240-285. https://doi.org/10.1086/284133
Šimčikas, S. (2019) Corporate campaigns affect 9 to 120 years of chicken life per dollar spenthttps://forum.effectivealtruism.org/posts/L5EZjjXKdNgcm253H/corporate-campaigns-affect-9-to-120-years-of-chicken-life
Seljetun, K. O., Eliassen, E., Madslien, K., Viljugrein, H., Vindenes, V., Øiestad, E. L., & Moe, L. (2019). Prevalence Of Anticoagulant Rodenticides In Feces Of Wild Red Foxes (Vulpes Vulpes) In Norway. Journal of Wildlife Diseases. In-Press. https://doi.org/10.7589/2019-01-027
Serieys, L. E., Bishop, J., Okes, N., Broadfield, J., Winterton, D. J., Poppenga, R. H., ... & O'Riain, M. J. (2019). Widespread anticoagulant poison exposure in predators in a rapidly growing South African city. Science of the Total Environment, 666, 581-590. https://doi.org/10.1016/j.scitotenv.2019.02.122
Slingerland, L. I., Fazilova, V. V., Plantinga, E. A., Kooistra, H. S., & Beynen, A. C. (2009). Indoor confinement and physical inactivity rather than the proportion of dry food are risk factors in the development of feline type 2 diabetes mellitus. The Veterinary Journal, 179(2), 247-253. https://doi.org/10.1016/j.tvjl.2007.08.035
Tarkosova, D., Story, M. M., Rand, J. S., & Svoboda, M. (2016). Feline obesity–prevalence, risk factors, pathogenesis, associated conditions and assessment: a review. Veterinární Medicína, 61(6), 295-307. https://www.agriculturejournals.cz/publicFiles/145_2015-VETMED.pdf
Themb’alilahlwa, A. M., Monadjem, A., McCleery, R., & Belmain, S. R. (2017). Domestic cats and dogs create a landscape of fear for pest rodents around rural homesteads. PloS one, 12(2), e0171593. https://doi.org/10.1371/journal.pone.0171593
Thomas, R. L., Fellowes, M. D., & Baker, P. J. (2012). Spatio-temporal variation in predation by urban domestic cats (Felis catus) and the acceptability of possible management actions in the UK. PloS one, 7(11), e49369. https://doi.org/10.1371/journal.pone.0049369
Tschanz, B., Hegglin, D., Gloor, S., & Bontadina, F. (2011). Hunters and non-hunters: skewed predation rate by domestic cats in a rural village. European Journal of Wildlife Research, 57(3), 597-602. https://doi.org/10.1007/s10344-010-0470-1
U.S. Environmental Protection Agency. (2006). Analysis of Rodenticide Bait Use. Memorandum from Biological Analysis Branch to Registration Branch. United States Environmental Protection Agency, Washington, D.C. http://fluoridealert.org/wp-content/pesticides/EPA-HQ-OPP-2006-0955-0004.pdf
van Heezik, Y., Smyth, A., Adams, A., & Gordon, J. (2010). Do domestic cats impose an unsustainable harvest on urban bird populations?. Biological Conservation, 143(1), 121-130. https://doi.org/10.1016/j.biocon.2009.09.013
Washington Post (2016). Got rats? These homeless cats are for hire. https://www.washingtonpost.com/news/animalia/wp/2016/05/06/got-rats-these-homeless-cats-are-for-hire/
Washington Post (2019). How many Americans have pets? An investigation of fuzzy statistics. Jan 19, 2019. https://www.washingtonpost.com/science/2019/01/31/how-many-americans-have-pets-an-investigation-into-fuzzy-statistics/
Willson, S. K., Okunlola, I. A., & Novak, J. A. (2015). Birds be safe: can a novel cat collar reduce avian mortality by domestic cats (Felis catus)?. Global Ecology and Conservation, 3, 359-366. https://doi.org/10.1016/j.gecco.2015.01.004
Wolf, P. J. (2016). What If Everything You Thought You Knew About “Feral” Cats Was Wrong?. In Proceedings of the Vertebrate Pest Conference (Vol. 27, No. 27). https://escholarship.org/uc/item/6ks593jz
Woods, M., McDonald, R. A., & Harris, S. (2003). Predation of wildlife by domestic cats Felis catus in Great Britain. Mammal review, 33(2), 174-188. https://doi.org/10.1046/j.1365-2907.2003.00017.x
Credits
![]()
This essay is a project of Rethink Priorities.
It was written by Kim Cuddington with contributions from Derek Foster and David Moss. Jane Capozzelli, Marcus Davis, Michelle Graham, Peter Hurford, Simon Eckerström Liedholm, Abraham Rowe, Jason Schukraft, Daniela R. Waldhorn, and Saulius Šimčikas provided helpful comments on this essay.
If you like our work, please consider subscribing to our newsletter. You can see all our work to date here.
.
I'll ask whoever runs the utility.farm site to update my piece cited in this with a note that the cost-effectiveness estimates might be based on bad estimates of cat impact.
Additionally, my cost-effectiveness estimates were only for the US - it is probably most cost-effective to work on cat predation in countries like the UK where a much higher percentage of outdoor cats are owned.
I find the comments about rodents/birds interesting, but mostly irrelevant to the discussion of cat predation, and find framing addressing cat predation and improving rodent welfare as competing aims very strange. I'm going to refer to rodents below, but it could apply to any animals killed by cats. It doesn't seem obvious that the causal chain we should care about is stopping cat predation causing painful rodent deaths; instead, we should consider both rodenticides causing painful rodent deaths and cat predation causing painful rodent deaths to be important issues.
For there to be a coherent argument to not address cat predation, you would need to demonstrate not only that rodenticides more painful than death via cats, but have a picture of the average rodent's life after the moment they might have been killed by a cat. Since any rodent who is killed by a cat definitionally would have lived longer had it been killed by a rodenticide instead, the rodent is going to accumulate further positive and negative experiences during its life before being killed. Even if rodenticides are twice as painful, it seems reasonable to expect a prolonged life to often be good, and outweigh that.
Regardless, this doesn't seem like an argument against addressing cat predation - its an argument that the most effective way to address rodent suffering might be both addressing cat predation and addressing painful rodenticides.
A human analogy might be: we shouldn't address malaria because someone dying from malaria, if saved, might die from another more painful disease later. I think what follows from that is that we should address malaria and the other painful disease. Not that we should let malaria kill the person / say that it is unclear if malaria is good or bad. Addressing malaria is clearly good, as is reducing cat predation. There might be unintended effects of both that also need to be addressed. But it doesn't mean that addressing those things has an unclear sign.
My point is, cat predation is clearly bad for rodents. Rodenticides are also clearly bad. We should probably address both things, and be aware of the effect of only addressing one, but not that we shouldn't address either. Broadly, this seems to apply to wild animal welfare issues in general - the downstream effects are really really complicated, but by doing monitoring during interventions to see unanticipated effects, and addressing those as they come up, we probably make more progress than just pointing out the complexity.
I guess the question that is raised is, how far down should we care about downstream effects from interventions, as opposed to just monitoring interventions and addressing effects as they arise.
The position of this piece is that, given we have no knowledge regarding the central tendency and distribution of rodent well-being, all we can do realistically is compare the quality of deaths. Whether you view an earlier death as a positive or a negative depends on both whether you assume rodent lives are on average net positive or net negative, and whether you assume the subsequent numerous offspring are likely to experience net positive or net negative lives. We are agnostic on this, but do point out that the length of time suffering due to poisoning or starvation will be longer than the suffering due to cat predation. However, one point we raised regarding outdoor cat presence is of course that fear-based vicinity control could reduce total deaths due to both poisoning and predation (although we certainly accept that this is this is not a simple approach Bedoya-Perez et al 2019, Krijger et al. 2017).
These points aside, the reason that we frame cat predation and rodenticide use as alternatives is that if you prioritize human interests and welfare above wildlife interests (and again we are agnostic here) then you will absolutely need to come up with an alternative method of control. Commensal rodent populations have large economic and human health impacts (e.g. Meerburg et al. 2009, Stenseth et al. 2003). (As an aside, given the tremendous reproductive output, you will also need controls if you consider rodent starvation something to be averted). With respect to downstream effects, cat predation and fear generation are methods of control that may be under appreciated and which at least don't have the far-reaching chemical contamination issues that rodenticides do.
Lit cited
Bedoya-Perez, M. A., Smith, K. L., Kevin, R. C., Luo, J. L., Crowther, M. S., & McGregor, I. S. (2019). Parameters that affect fear responses in rodents and how to use them for management. Frontiers in Ecology and Evolution, 7, 136. https://doi.org/10.3389/fevo.2019.00136
Krijger, I. M., Belmain, S. R., Singleton, G. R., Groot Koerkamp, P. W., & Meerburg, B. G. (2017). The need to implement the landscape of fear within rodent pest management strategies. Pest management science, 73(12), 2397-2402. https://doi.org/10.1002/ps.4626
Meerburg, B. G., Singleton, G. R., & Kijlstra, A. (2009). Rodent-borne diseases and their risks for public health. Critical reviews in microbiology, 35(3), 221-270. https://doi.org/10.1080/10408410902989837
Stenseth, N. C., Leirs, H., Skonhoft, A., Davis, S. A., Pech, R. P., Andreassen, H. P., ... & Zhang, Z. (2003). Mice, rats, and people: the bio‐economics of agricultural rodent pests. Frontiers in Ecology and the Environment, 1(7), 367-375. https://doi.org/10.1890/1540-9295(2003)001[0367:MRAPTB]2.0.CO;2
Thanks for the detailed response. I think I disagree in a sort of principled way with particular kinds of approaches to downstream effects, in part because I think it could just turn into an endless game of trying to figure out how things could turn out poorly, as opposed to a model where we address both rodenticides and cat predation (though I recognize I am stubbornly resisting you all trying to do prioritization, which might not be a good idea given the name of your organization).
Regardless, I'm drafting a new intro section for my cost-effectiveness updates linking to your updated numbers. Thanks again for doing the analysis!
Interesting. If rodent and bird mortality from cats is generally compensatory, then perhaps the collar-based deterrents has benefits by allowing animals in good health time to escape while also allowing for predation of individuals in the "doomed surplus." Just read some papers on the idea that low levels of predation has benefits to population growth (e.g. Sandercock et al. 2011), and am now curious as to whether individuals, on average, would be "functioning and feeling better" commensurate with change in population size... unknown.
Sandercock BK, Nilsen EB, Broseth H, Pedersen HC (2011) Is hunting mortality additive or compensatory with natural morality? Effects of experimental harvest on the survival and cause-specific mortality of willow ptarmigan. Journal of Animal Ecology 80:244-258. doi: 10.1111/j.1365-2656.2010.01769.x
Very interesting... particularly this point "The impacts of harvest on seasonal and annual variation in survival likely differ among populations and animal species exposed to different sets of environmental conditions". That is, whether mortality is compensatory or additive depends on context (as always.... ecology is just not easy to generalize).
I'm also really curious as to number of natural predators in areas were we expect high outdoor cat presence (e.g., low density housing) and how that plays out re compensatory and additive mortality.
your appendix certainly highlights how spatially-simple the Loss et al. 2013 estimate of cat-caused mortality is.... seems like their model could be improved by adding in a metric of urban landcover (e.g. LANDSAT impervious surface).... perhaps hotspots could be highlighted if compared against eBird and/or Partners In Flight data on avian abundance.... I similarly agree, as you allude to, there appear to be opportunities to use urban landcover as an approximation of predator-prey dynamics (via trophic structure, perhaps), but also pitfalls given hyper-local contexts. thanks for the opportunity to think out loud here ..... :)